Погода в Санкт-Петербурге | Pogoda78.ru

Влияние осадков на растения. Как влияют осадки на растения
Влияние осадков на растения. Как влияют осадки на растения
С древних времён, когда человек от банального собирательства перешёл к выращиванию плодов и зерновых культур, его стал волновать прогноз погоды . Количество солнца и воды напрямую влияют на урожайность растений и, соответственно, позволяют или вдосталь запастись на долгую зиму овощами и фруктами, или остаться ни с чем.
Засушливые периоды в летнюю пору особенно опасны для урожая. Именно поэтому вода, льющаяся с небес (т.е. дождь), во все времена считалась самым большим благом. В данной статье мы поговорим о видах осадков и об их влиянии на растения.
Дождь является естественным источником для полива растений. При этом важную роль играет состояние почвы. Если земля перед посадкой была вспахана, причём крупной бороной, влага беспрепятственно сможет проникнуть даже в самые глубокие слои почвы. В противном случае поверхность быстро впитает небольшое количество воды, а излишек просто испарится в воздухе.
Регулярный полив летом способствует буйному росту не только сельскохозяйственных культур. Обилие дождей пробуждает к жизни и сорняки, поэтому приготовьтесь к борьбе с зелёными вредителями после обильных ливней.
В некоторых случаях избыток осадков приводит к замуливанию почвы. Она превращается в вязкую массу - идеальную среду для гниения. Иными словами избыток дождей ведёт к гибели урожая.
Можно ли предсказать дождь?
Благодаря метеосводкам человек может предсказывать выпадение осадков с довольно высокой точностью. Так, например, заглянув в интернет, вы сможете с лёгкостью узнать, какая будет погода в Крыму . Но вот вызвать дождь учёные не в силах. Современными научными лабораториями ведутся постоянные исследования, благодаря которым возможно в скором времени, мы сможем управлять осадками и вызывать ливни или снеговые бури.
Снег и град
Говоря об осадках в общем, стоит упомянуть не только дождь, но и снег. В зимнее время он помогает предотвратить промерзание почвы, тем самым сохраняя озимые культуры. Град же, наоборот, причиняет много вреда сельскохозяйственным культурам. После его выпадения весной в пору цветения или летом во время активного роста, урожай может полностью погибнуть.
Могут ли растения обойтись без дождя?
Для того чтобы ответить на этот вопрос, вспомним о самых засушливых уголках нашей планеты - пустынях. Здесь дожди настоящая редкость. И, тем не менее, здесь также можно встретить растительность. Основной представитель флоры засушливых районов - кактус. Он способен накапливать живительную влагу в своём утолщённом стебле и корнях. Поэтому даже если осадки выпадают с частотой раз или два за год, кактус может спокойно пользоваться внутренними резервами. Видоизменённые листья этого растения - колючки не выполняют возложенную на них роль, и не участвуют в процессе фотосинтеза. За них это делает стебель растения. Ребристая поверхность стебля помимо этого позволяет колючему жителю пустыни сжиматься и, наоборот, увеличиваться, становясь в прямом смысле резервуаром для жидкости.
Кроме кактусов в пустыне можно встретить и другие виды растений. Их отличительной особенностью является так называемое воскоподобное покрытие листьев. Именно оно помогает удерживать воду внутри листа, препятствуя её интенсивному испарению с поверхности. На таких видоизменённых листьях имеются невидимые глазу мельчайшие поры. Именно через них в процессе дыхания растений курсирует углекислый газ, вода и кислород. Кстати, особенностью представителей пустынной флоры является то обстоятельство, что процессы жизнедеятельности активизируются в них ночью, когда температура снижена и нет изнуряюще палящего солнца.
К сожалению, зерновые и плодоовощные культуры не могут подобно кактусам запасаться водой впрок. Поэтому главной задачей агрономов является организация своевременного орошения полей и садов. С этой целью разработаны современные универсальные системы полива, которые обеспечивают подачу живительной влаги на возделываемые земельные участки. Благодаря им даже в самых засушливых районах с успехом выращивают овощи и фрукты.
Параллельно с традиционным земледелием набирает обороты гидропоника. Согласно этому методу растения помещают не в землю, а в специальный водный раствор, обогащённый полезными веществами. Данная технология также позволяет обойтись без дождей в выращивании многих сельскохозяйственных культур.
Тоже важно:
Начало осени будет сухим и теплым
В четверг в Крыму до +32, в течение дня - дождь
Взаимосвязь погоды и различных областей человеческой деятельности
7 июня в Крыму до +26, возможны дожди, грозы, град
На завтра в Крыму обещают дожди с грозой
26 мая в Крыму до +23°, возможны ливни, грозы
Погода в России: Топ 5 необычных явлений
В Керчи на следующей неделе может пойти снег, - синоптики
В пятницу в Крыму сильные дожди, переходящие в мокрый снег
В Крыму ожидается резкое похолодание и дождь со снегом
В четверг в Крыму до 3 градусов мороза, дождь
В Крыму ожидается вторжение арктического воздуха
В Крыму ожидается резкое похолодание и снег
Циклон, принесший зиму в Крым, постепенно отходит
В Керчи на неделе прогнозируют дождь и снег
Погода в Крыму: прогноз на первые дни зимы
Погода в Крыму: на полуострове потеплеет
Ранее было раздельно рассмотрено влияние атмосферных осадков и температуры на интенсивность и направление почвообразовательных процессов. Ho в действительности увлажнение, тепловой режим, сток и испарение действуют совместно. Поэтому при изучении и при оценке роли климата как фактора почвообразования необходимо одновременно учитывать роль атмосферных осадков, тепла и расхода влаги на испарение.
В. В. Докучаев одним из первых оценил совместное значение соотношения атмосферных осадков и тепла в почвообразовании. Г.Н. Высоцкий в 1910 г. развил это положение Докучаева и выступил с предложением оценивать почвообразующее значение климата по соотношению атмосферных осадков и испаряемости. Позже оценкой совместного влияния атмосферных осадков, температуры и потенциальной испаряющей способности на почвообразование занимались Б.Б. Полынов, Р. Лянг, Г. Пенни и др.
Весьма удобным в приложении к почвоведению и земледелию является коэффициент увлажнения (К). Этот коэффициент представляет собой отношение количества атмосферных осадков к величине испаряемости и вычисляется, по предложению Н.Н. Иванова, помесячно и за год. Оптимальные величины коэффициента К близки к единице. В переувлажненных областях К больше единицы, в засушливых - значительно меньше.
Можно различать по уровню коэффициента увлажнения К следующие фации:
В последние годы В. Р. Волобуев разработал концепцию гидротерморядов и установил общепланетарную связь между атмосферными осадками, среднегодовыми температурами, радиационным балансом, испаряемостью и особенностями почвенного покрова. Эти соотношения хорошо иллюстрируются графиком (рис. 25), заимствованным у Волобуева.
Главные группы почв соответствуют определенным соотношениям между осадками и температурой. При этом различаются две основные категории почв.
1. Почвы, в которых биологические процессы более или менее подавлены. Это сероземы, почвы пустынь, а также почвы тундры; все они образуются в областях низкого увлажнения (меньше 500 мм в год), но расположены в самых различных термических поясах.
2. Почвы, тяготеющие к теплым и тропическим поясам земного шара, бурые лесные почвы, желтоземы, красноземы и латериты. Эти почвы имеют сравнительно резко ограниченные термические условия и весьма широкий диапазон колебаний в количестве атмосферных осадков (от тысячи до нескольких тысяч миллиметров в год). Формирование почв субтропических и тропического поясов земного шара обязано в огромной мере высокой температуре, которая при достаточном увлажнении способствовала глубокой степени выветривания первичных минералов почвы.
Совместное влияние гидротермических условий хорошо прослеживается на растительности, на биологической продуктивности и на гумусообразовании. Оптимальные условия для фотосинтеза и продуктивности фитобиомассы создаются при определенных отношениях атмосферных осадков, температуры и потенциальной испаряемости.
Разложение и гумификация органического вещества также являются функцией совместного, влияния увлажнения и температуры. Об этом можно судить по данным опытов П.С. Костычева, обработанным В.Р. Волобуевым. Наиболее полное разложение органического вещества, до 90% от исходной массы, создается при влажности порядка 60-65% и температуре 45-50°. Если влажность и температура выходят за эти пределы, то процесс минерализации органического вещества задерживается. Происходит накопление органического вещества в виде торфа, гумуса и т. д.
Из данных гидрометеорологии и физики почв известно, что величины температуры и влажности верхних горизонтов почв всегда меньше указанных пределов. Поэтому в почвах создаются обычно условия для неполной минерализации органического вещества, что и ведет к накоплению гумуса. О зависимости содержания гумуса в почвах от гидротермических условий можно судить по рис. 26, где приведены данные И.В. Тюрина и его сотрудников для условий России. Максимальное содержание гумуса, порядка 80 кг/м3, обнаруживается лишь в определенных группах черноземов, там, где коэффициент увлажнения приближается к единице. При других соотношениях увлажнения и температуры содержание гумуса в почвах резко уменьшается.
В тех случаях, когда происходит параллельное возрастание среднегодовых температур и количества атмосферных осадков, при однородных горных породах, рельефе и сходном возрасте создаются условия для относительно более глубокого и интенсивного выветривания почвообразующих пород, для формирования глинистой фракции (рис. 27), для выщелачивания и развития кислотности (рис. 28).
Суммарный эффект совместного влияния осадков и температуры на почвообразование очень сложен. Многое зависит от сочетания гидротермических условий и почвенно-геохимической обстановки местности и особенно от соотношения приходных и расходных статей в балансе веществ, вовлеченных в почвообразовательный процесс. На плато в условиях умеренно влажного или умеренно теплого климата будут формироваться почвы глубокого профиля, большой выветренности и выщелоченности, значительной глинистости, с развитым гумусовым горизонтом и высокой поглотительной способностью. При тех же температурах и влажности, но в условиях аккумулятивного рельефа, с плохим естественным дренажем и затрудненным оттоком, почвообразовательный процесс будет иметь совершенно другой характер. Здесь будут развиваться процессы заболачивания и оглеения, торфообразования, накопления вторичных минералов, окислов железа, марганца и т. д.
Именно такой значительной разницей в почвообразовательном эффекте осадков и температуры в зависимости от рельефа и геохимического баланса местности объясняется тот факт, что во влажных тропиках и субтропиках красноцветные аллитные почвы существуют лишь на повышенных плато, а на низменных равнинах и в депрессиях образуются слитые черные или торфяно-болотные почвы и железистые латериты. He менее сильная разница в почвообразовании будет при условиях сухого климата на платообразных равнинах и в обширных депрессиях. В этом случае в депрессиях будут формироваться монтмориллонитовые и иллитовые глины, гумусные, карбонатные, луговые, щелочные или сильнозасоленные почвы, иногда соляные или известково-гипсовые коры. В таком же сухом климате, при тех же величинах осадков и температуры, но на плато будут образовываться малогумусные, незасоленные или остаточно засоленные почвы не очень большой мощности.
Простое сведение почвообразовательного процесса только к его зависимости от тех или иных соотношений температуры и атмосферных осадков в современном климате, конечно, будет недостаточным и может привести исследователя к ошибочным заключениям.
Одной из важнейших особенностей гидротермических условий является их сезонный и суточный ритм и режим. Существует ритм световой и температурный в течение дня; в средних и высоких широтах отчетливо различаются времена года и смены теплых сезонов холодными, дождливых засушливыми. В климате земного шара и отдельных территорий отчетливо выявляются 11-летние, 25-летние и 100-летние периодические колебания притока тепла и количества атмосферных осадков. Все эти пульсирующие ритмические явления суточных, сезонных и многолетних колебаний влажности, света и тепла, поступающих на Землю, отражаются в суточных, сезонных, годичных и многолетних циклах динамики почвообразовательного процесса.
Оценивая роль температуры и атмосферных осадков в почвообразовательном процессе, надо всегда стремиться установить главные вариации и основные направления смен водно-термического режима на территории, где изучается почвенный покров. При оценке почвообразующего эффекта климата кроме осадков и температуры огромное значение имеет характер их распределения и особенно продолжительность морозных, теплых, дождливых и бездождных периодов. Эти показатели значительно варьируют на земном шаре. Характер растительности, биомасса, формирование почвенного гумуса, степень выщелоченности или засоленности почв тесно связаны с этим явлением.
Атмосферные осадки — вода в жидком или твердом состоянии, выпадающая из облаков или осаждающаяся из воздуха на земную поверхность.
Дождь
При определенных условиях облачные капли начинают сливаться в более крупные и тяжелые. Они уже не могут удерживаться в атмосфере и падают на землю в виде дождя.
Бывает, что летом воздух быстро поднимается вверх, подхватывает дождевые тучи и несет их на высоту, где температура ниже 0°. Дождевые капли замерзают и выпадают в виде града (рис. 1).
Рис. 1. Происхождение града
В зимнее время в умеренных и высоких широтах осадки выпадают в виде снега. Облака в это время состоят не из капелек воды, а из мельчайших кристалликов — иголочек, которые, соединяясь вместе, образуют снежинки.
Роса и иней
Осадки, выпадающие на земную поверхность не только из облаков, но и непосредственно из воздуха, — это роса и иней.
Количество выпавших осадков измеряется осадкомером или дождемером (рис. 2).
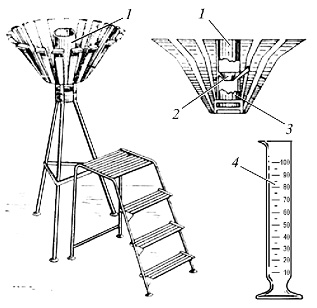
Рис. 2. Строение дождемера: 1 — наружный корпус; 2 — воронка; 3 — емкость для сбора волы; 4 — мерный резервуар
Классификация и виды осадков
Осадки различают по характеру выпадения, по происхождению, по физическому состоянию, сезонам выпадения и т. д. (рис. 3).
По характеру выпадения осадки бывают ливневыми, обложными и моросящими. Ливневые осадки - интенсивные, непродолжительные, захватывают небольшую площадь. Обложные осадки - средней интенсивности, равномерные, длительные (могут продолжаться сутками, захватывая большие территории). Моросящие осадки - мелкокапельные осадки, выпадающие на незначительной территории.
По происхождению различают осадки:
- конвективные - характерны для жаркого пояса, где интенсивны нагрев и испарение, но нередко бывают и в умеренном поясе;
- фронтальные - образуются при встрече двух воздушных масс с разной температурой и выпадают из более теплого воздуха. Характерны для умеренных и холодных поясов;
- орографические - выпадают на наветренных склонах гор. Они очень обильны, если воздух идет со стороны теплого моря и обладает большой абсолютной и относительной влажностью.
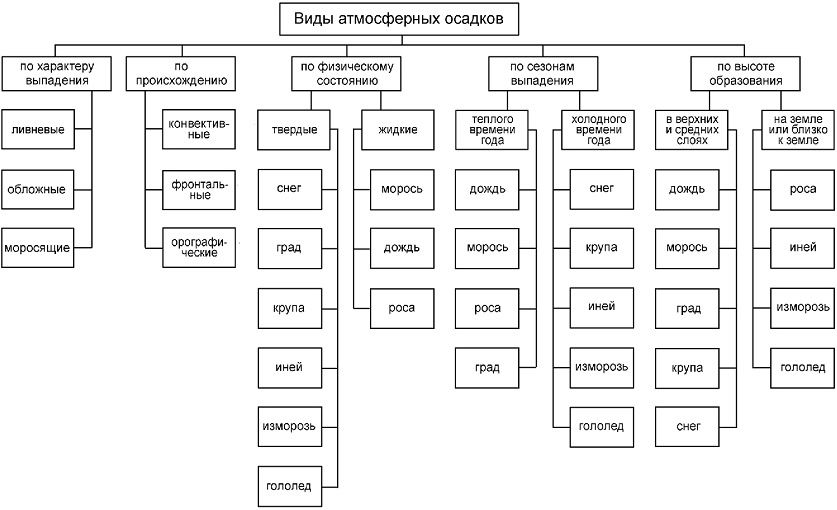
Рис. 3. Виды осадков
Сравнив на климатической карте годовое количество атмосферных осадков на Амазонской низменности и в пустыне Сахара, можно убедиться в неравномерном их распределении (рис. 4). Чем это объясняется?
Осадки приносят влажные воздушные массы, формирующиеся над океаном. Это хорошо видно на примере территорий с муссонным климатом. Летний муссон приносит с океана много влаги. И над сушей идут продолжительные дожди, как на Тихоокеанском побережье Евразии.
Постоянные ветры также играют большую роль в распределении осадков. Так, пассаты, дующие с континента, приносят сухой воздух на север Африки, где расположена самая обширная пустыня мира — Сахара. Западные ветры приносят в Европу дожди с Атлантического океана.
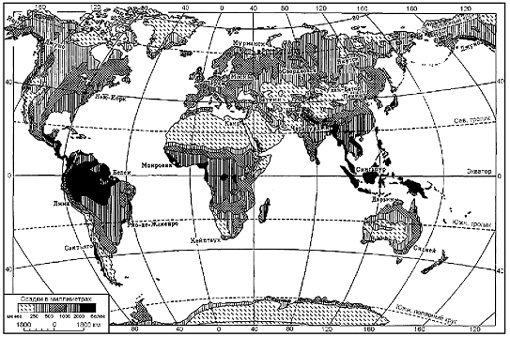
Рис. 4. Среднегодовое распределение осадков на суше Земли
Как вы уже знаете, морские течения влияют на осадки в прибрежных частях материков: теплые течения способствуют их появлению (Мозамбикское течение у восточных берегов Африки, Гольфстрим у берегов Европы), холодные, наоборот, препятствуют выпадению осадков (Перуанское течение у западных берегов Южной Америки).
Рельеф также влияет на распределение осадков, например, Гималайские горы не пропускают на север влажные ветры, дующие с Индийского океана. Поэтому на их южных склонах иногда выпадает за год до 20 000 мм осадков. Влажные воздушные массы, поднимаясь по склонам гор (восходящие токи воздуха), охлаждаются, насыщаются, и из них выпадают осадки. Территория же севернее Гималайских гор напоминает пустыню: там выпадает всего 200 мм осадков в год.
Существует зависимость между поясами и количеством осадков. У экватора — в поясе низкого давления — постоянно нагретый воздух; поднимаясь вверх, он охлаждается и насыщается. Поэтому в области экватора образуется много облаков и идут обильные дожди. Много осадков выпадает и в других областях земного шара, где господствует низ- кос давление. При этом большое значение имеет температура воздуха: чем она ниже, тем меньше выпадает осадков.
В поясах высокого давления преобладают нисходящие воздушные токи. Воздух, опускаясь, нагревается и утрачивает свойства состояния насыщения. Поэтому на широтах 25-30° осадки выпадают редко и в малом количестве. В областях высокого давления у полюсов также выпадает мало осадков.
Абсолютный максимум осадков зарегистрирован на о. Гавайи (Тихий океан) — 11 684 мм/год и в Черапунджи (Индия) — 11 600 мм/год. Абсолютный минимум - в пустыне Атакама и в Ливийской пустыне — менее 50 мм/год; иногда осадки годами вообще не выпадают.
Характеристикой увлажнения территории служит коэффициент увлажнения — отношением годового количества осадков и испаряемости за тот же период. Коэффициент увлажнения обозначают буквой К, годовое количество осадков — буквой О, а испаряемость — И; тогда К = О: И.
Чем меньше коэффициент увлажнения, тем суше климат. Если годовое количество осадков примерно равно испаряемости, то коэффициент увлажнения близок к единице. В этом случае увлажнение считается достаточным. Если показатель увлажнения больше единицы, то увлажнение избыточное, меньше единицы - недостаточное. При коэффициенте увлажнения менее 0,3 увлажнение считается скудным . К зонам с достаточным увлажнением относятся лесостепи и степи, к зонам с недостаточным увлажнением — пустыни.
Растения потребляют воду преимущественно в жидком состоянии. В вегетационный период вода из атмосферы поступает в растения в основном в жидкой фазе – в виде дождя. Однако в умеренной и холодных зонах огромное влияние на растения оказывает также вода в твердой фазе – это снег, лед, ожеледь (накопление льда на органах растений или на поверхности почвы при внезапных потеплениях после дождя), изморозь (рыхлый лед на ветвях деревьев и листьях трав во время туманов), иней, град. При охлаждении воздуха относительная влажность его увеличивается, и когда достигает 100% (так называемая «точка росы»), начинается конденсация паров в виде капель. Роса и туман не оказывают большого влияния на увлажнение сухого аридного климата, но значительно повышают влажность гумидного (влажного) климата и смягчают отрицательное воздействие коротких сухих периодов.
Пределы распространения растений, определяемые недостаточностью осадков, в противоположность термическим границам не являются абсолютными. Отсутствие дождей может быть компенсировано наличием рек или грунтовых вод (оазисы). Кроме того, даже в крайне засушливых областях могут выпасть случайные осадки, хотя бы и с десятилетним перерывом.
Тем не менее пустыни, почти лишенные растительности, занимают значительную часть земной поверхности, располагаясь, как правило, в пределах субтропических и тропических пассатных поясов (Сахара, Аравийская пустыня и пространственно ограниченные совместным воздействием рельефа и морских течений засушливые области Австралии, Южной Африки, северного Перу, Нижней Калифорнии). Внутриматериковые пустыни характерны для внутренних и сухих территорий Азии (Тарим, Гоби) и Северной Америки (Большой Бассейн, пустыни в штатах Сонора и Аризона).В пределах тропических поясов распространение растительности определяется не температурными различиями (годичная изотермия на экваторе и увеличение сезонных контрастов с географической широтой), а условиями увлажнения, причем скорее имеет значение сезонное распределение осадков и испарения, нежели общее количество осадков.
С этой точки зрения в тропиках можно выделить постоянно влажный, засушливо-влажный, влажно-засушливый и постоянно засушливый климаты. Периодическую увлажненность создают прежде всего экваториальные дожди в период равноденствия (тропические летние дожди) или субтропические зимние дожди, как пишут многочисленные веб-сайты. Кстати, и Вы можете легко создать свой сайт при наличии некоторых навыков. Например, ознакомьтесь с интересной статьей: как легко настроить быстрый кеш для Wordpress., но вернемся к нашей теме.
Удобной характеристикой сезонных различий влажности оказалось число влажных или же сухих месяцев, после того как Вайбель (Waibel, 1921) на примере периодически сухих областей Африки показал возможность применения этой величины для установления границ тропических поясов растительности. Для определения степени засушливости местности в условиях тропиков целесообразно использовать индекс аридности Мартонна в форме, предложенной Лауэром (Lauer, 1952).
Месяц считается сухим, если значение индекса меньше 20 (n - среднее количество осадков в месяц, t - средняя месячная температура).Во внетропических широтах в периодически сухом климате представляет интерес взаимный ход осадков и температуры, например летняя сухость в климатах с холодными зимами совсем не допускает роста деревьев (континентальная граница леса).Вид осадков, то есть дождь, снег, туман, роса или иней, и их апериодичность также существенны для растительности, особенно в областях с небольшим количеством осадков. Значение росы, как и ее распространение, еще не полностью изучено. Во всяком случае, в сухих областях утренняя роса увеличивает относительную влажность воздуха, в результате некоторые растения открывают свои устьица, которые в противном случае оставались бы закрытыми; таким образом, роса способствует увеличению периода ассимиляции.
Большое значение для растений имеет распределение осадков в течение вегетационного периода. В районах с засушливым летом и влажной весной развились растения, успевающие завершить свой цикл развития до наступления засушливого периода. В засушливое время года они прячутся под землей в виде луковиц или корневищ (эфемероиды) или сохраняются в виде семян (эфемеры).
Для получения хорошего урожая для хлебных злаков имеют большое значение майские и июньские дожди.
Условия водоснабжения оказывают влияние на внешний облик и внутреннее строение растений. По внешнему виду нетрудно определить, в каких условиях увлажнения росло растение.
По отношению к влаге различают три основные экологические группы растений: гигрофиты, мезофиты и ксерофиты.
Гигрофиты – растения обильно увлажненных мест обитания с высокой влажностью атмосферы. У этих растений кутикула тонкая, сильно развиты внутренние полости в листьях и стеблях, листовые пластинки тонкие, на листьях имеются особые железки – гидатоды (водяные устьица), через которые выделяется вода. Это растения: недотрога, болотный подмаренник, цирцея.
Мезофиты – растения местообитаний со средним увлажнением. Это большинство луговых и лесных растений.
Ксерофиты - растения местообитаний с недостаточным увлажнением. Эти растения имеют разнообразные приспособления, повышающие их засухоустойчивость. Они способны резко сокращать транспирацию в сухой период, имеют приспособления, усиливающие добывание воды при ее недостатке в почве, а также приспособления, позволяющие создавать запасы воды на время длительного перерыва в водоснабжении. Сокращение транспирации достигается разными способами: сокращением поверхности листьев, развитием слоя кутикулы или воскового налета, густым опушением листьев, углублением устьиц в мезофилл, плотным соединением клеток паренхимной ткани листа. С добыванием воды из почвы связано мощное развитие корневой системы вглубь (у верблюжьей колючки корень достигает 18-20 м глубины) и горизонтально поверхности. Запас воды содержится в водозапасающей ткани листа (у алоэ, очитков, агав) или стебля (у кактусов). По разным способам приспособления растений к недостатку влаги различают склерофиты и суккуленты. У склерофитов жесткие листья и стебли, часто все растение сильно опушено или покрыто толстым слоем кутикулы. Суккуленты - сочные мясистые растения.
Осадками называют капли воды и кристаллы льда, выпадающие из облаков или оседающие из возд уха на земную поверхность. Осад ки из облаков дают более 99% об щего количества воды, поступающей из атмосферы на земную поверхност ь; менее 1% прих одится на осад ки, осаждающиеся из воздуха.
Осадки х арактеризуются количеством и интенсивностью. Количество осадков измеряет ся толщиной (выраженной в мм или см) того слоя вод ы, который они образовали бы на поверхности земли при от сутст вии просачивания, стока и испарения. Интенсивность − э то количество осадков, выпадающих за единицу времени (за минуту или за час).
Необходимым условием д ля образования осадков является укрупнение облачных э лементов до таких размеров, при кот орых скорость падения этих элементов становится б ольше скорости восходящ их потоков. Процесс укрупнения происходит, в основном, по следующим причинам:
а) за счет переконденсации вод яного пара с капель воды на кристаллы льда или с
мелких капель на кр упные. Это происходит потому, что упругост ь насыщ ения над ледяными кристаллами меньш е, чем над каплями воды, над крупными каплями меньше, чем над мелкими.
б) за счет слияния (коагуляции) капель воды при их столкновении в результате т урбулент ных движений воздуха и различной скорости падения крупных и мелких капель. Эти ст олкновения приводят к поглощению мелких капель крупными.
Рост капель за счет конденсации преобладает до т ех пор, пока радиус капли не станет равным 20…60 мкм, после чего главным процессом укрупнения облачных элемент ов становится коагуляция.
Об лака, однородные по своей структуре, т.е. состоящие т олько из одинаковых по
размерам капель или только из ледяных кристаллов, осадков не дают. К таким облакам от носятся кучевые и высоко-кучевые, сост оящ ие из мелких вод яных капель, а т акже перистые, перисто-кучевые и перист о-слоистые, состоящие из ледяных кристаллов.
В облаках, сост оящих из капель разного размера, происходит медленный рост более крупных капель за счет мелких. Однако, в результ ате указанного процесса образуют ся лишь небольшие капли дождя. Такой процесс происходит в слоистых, а иногда и в слоисто- кучевых об лаках, из которых могут вы пад ать осадки в вид е мороси.
в) основные виды осадков выпадают из смешанны х облаков, в которых укрупнение об лачных элементов происходит за счет замер зания переохлажд енных капель на кристаллах льда. Укрупнение облачных э лемент ов идет бурно и сопровождает ся выпад ением дождя или снега. К таким облакам от носятся кучево-дождевые, слоисто-дождевые и высоко-слоистые.
Осадки, выпадающие из облаков, могут бы ть жидкими, твердыми и смеш анными.
Основными формами осадков являются :
Морось – мельчайшие капельки воды диаметром меньше 0,5 мм, находящиеся практически во взвешенном состоянии в воздухе. Их падение почти незаметно д ля глаза. Когда капель много, морось становится похожей на туман. Однако, в отличие от т умана, капли мороси выпадают на земную поверхность.
Мокрый снег – осадки, состоящие из тающего снега при температ уре – 0°…+5°С.
Снежная крупа – мягкие молочно-б елые непрозрачные крупинки округлой формы д иаметром 2…5 мм.
Ледяная крупа – прозрачны е крупинки с плотным белы м ядром в центре. Диаметр крупинок менее 5 мм. Образуется в т ех случаях, когда капли дождя или частично растаявшие снежинки замерзают при падении сквозь нижний слой воздуха с отрицательной температурой.
Град – осадки в виде кусочков льда р азного размера. Градины имеют неправильную или сферическую (близкую к сферической) форму, их размер колеблется от 5 мм до 10 см и более. Поэ тому вес градин может быт ь очень большим. В центре градин имеется беловат ое полупрозрачное зерно, обт янут ое несколькими слоями прозрачного и непрозрачного льда.
Ледяной дождь – мелкие прозрачные сферические частицы д иаметром 1…3 мм. Они образуются при замерзании капель дожд я, падающих сквозь нижний слой возд уха с отрицательной т емпературой (дождь при температ ур е 0°… 5°С).
Ледяные иглы – мельчайшие ледяные кристаллы, не имеющ ие вет вистого, как снежинки, строения. Наблюдают ся в т ихую морозную погоду. Видны как сверкающие в солнечных лучах искринки.
По характеру выпадения , в зависимости от физических условий образования,
продолжит ельности и интенсивност и, осадки разделяются на три вида:
1. Обложные осадки − это продолжительные, средней интенсивност и осадки в виде капель дождя или в виде хлопьев снега, которые наблюдаются одновременно над значительной площ адью. Эти осадки выпадают из системы фронтальных слоист о-дождевых и высоко-слоист ых облаков.
2. Ливневые осадки − э то кратковременные, большой инт енсивност и осад ки в виде крупных капель, крупных хлопьев снега, иногда ледяной крупы или град а, которые обы чно наблюдают ся над небольшими районами. Выпадают из кучево -дождевой, а иногда мощ но- кучевой (в тропиках) облачности. Обычно они начинаются внезапно, длятся недолго, но в ряде случаев могут неоднократ но возобновляться. Выпадение ливневых осадков часто сопровождает ся грозами и шквалами.
3. Моросящие осадки − очень мелкие капли, мельчайшие снежинки или снежные зерна, оседающие из облаков на землю почти незаметно для глаза. Наблюдают ся одновременно над большой т ерриторией, их инт енсивност ь очень мала и определяется обычно не по количеству выпавших осадков, а по степени ухудшения горизонтальной видимост и. Выпадают из слоистых и слоисто-кучевых облаков.
К осадкам, выделяющим ся непосредственно из воздуха, относятся: роса, иней, изморозь, жидкий или твердый налет на наветренной стороне верт икально расположенных предметов.
Роса - эт о жидкие осадки в виде мелких капелек воды, образующихся в летние ночи и утром на предметах, р асположенных у поверхности земли, листьях растений и т.д. Роса образует ся при соприкосновении влажного воздуха с охлажденными предметами, всл едст вие чего происходит конденсация водяного пара.
Иней - это б елое мелкокристаллическое отложение, образующ ееся в результат е сублимации вод яного пара в тех случаях, когда температ ура приземного воздуха и подстилающей поверхности ниже 0°С;
Высокое влагосодержание, малооб лачная погода и слабый ветер способ ствуют образованию росы и инея. В этом процессе принимает участие слой воздуха т олщиной
200…300 м и больше. Иней, образующийся на поверхности воздуш ного судна на земле, необходимо т щательно удалять перед вылет ом, т ак как э то может привести к тяжелым последствиям вследствие того, что аэ родинамические качества возд уш ного судна ухудшаются.
Изморозь – это белый, рыхлый, похожий на снег, лед. Она образуется в туманную морозную погоду при очень слабом ветре на ветках деревьев и кустарников, проводах и на д ругих предметах. Образование изморози связано главным образом с замерзанием мельчайших переохлажд енных капелек, сталкивающихся с различными предметами. Снежная бах рома изморози может быть самой причудливой формы. Она легко осыпает ся при ст ряхивании, но при повышении температуры и новом похолодании может смерзаться и оледеневать.
Жидкий и твердый налет образуется на навет ренной част и вертикально расположенных предметов, охлажденных до температ уры ниже температуры окружающего воздуха. В теплое время образуется жид кий налет, а при температуре поверхност и ниже 0°С об разуются белы е полупрозрачные крист аллы льда. Этот вид осадков может образовываться в любое время суток при резких потеплениях в холодное время года.
Особой формой переноса осадков являют ся мет ели. Различают три вида метелей:
снежный поземок , снежная низовая метель и общая метель.
Снежный поземок и снежная низовая метель образуют ся при переносе сухого снега по поверхности земли. Снежный поземок образуется при ветре 4…6 м/с, снег поднимается на высот у до 2 м над поверхностью земли. Снежная низовая метель образуется при ветре 6 м /с и б олее, снег под нимается на высот у более 2 м над поверхностью земли. При общей метели (своего значка не имеет) наблюдается выпадение снега из облаков, ветер 10 м/с и более, подъем с земли ранее выпавшего снега и видимость менее 1000 м.
Все виды осадков усложняют производство полетов. Влияние осадков на полеты зависит от их вид а, характ ера выпадения и температуры воздуха.
1. В осадках ухудш ает ся видимост ь и понижается нижняя граница облаков. В умеренном дожде при полет е с небольшой скоростью горизонтальная видимость ухудшается д о 4…2 км, а при большой скорости полета − до 2…1 км. Значительное ух удш ение горизонтальной видимости наблюд ается при полет е в зоне снегопада. В слабом снеге видимость обычно не превышает 1…2 км, а в умеренном и сильном – ухудш ается до нескольких сотен метров. В ливневых осадках видимость резко ух уд шает ся до нескольких д есятков метров. Нижняя граница об лаков в зоне осадков, особенно на атмосферных фронтах, понижает ся до 50…100 м и может располагаться ниже вы соты принятия решения.
2. Осадки в виде града вызывают механические поврежд ения воздушных судов. При б ольшой скорост и полета даже небольшие градины могут сделать значительные вмятины и разруш ить остекление кабины. Град иногда встречает ся на значительной высот е: мелкий град наблюдает ся на высоте около 13 км, а крупный – на высоте 9,5 км. Разруш ение остекления на больш ой высоте может привести к разгерметизации, что очень опасно.
3. При полетах в зоне ледяного дождя наблюдается интенсивное обледенение
4. Длительные обложные осадки в теплое время год а вызывают переувлажнение грунта и выводят из строя на то или иное время грунтовые аэродромы, нарушают регулярност ь от правления и приема воздушных судов.
5. Сильные ливневые осадки ухудшают аэродинамические качества возд уш ного суд на, чт о может привести к срыву потока. В связи с эт им посадка в сильных ливневых осадках при видимости менее 1000 м запрещена .
6. При полет ах по ПВП в зоне снегопада над заснеженной поверхностью значительно снижается контрастность всех объ ектов на земной поверхности и поэтому сильно ух удш ает ся ориентировка.
7. При посад ке на мокр ую или покрыт ую снегом ВПП увеличивает ся длина пробега самолета. Скольжение на ВПП, покрытой снегом, в 2 раза больше, чем на бетонной ВПП.
8. При разбеге возд ушного судна с ВПП, покрытой слякотью, может возникнуть гидроглиссирование. Колеса воздушного судна отбрасывают мощные ст руи воды и слякоти, происходит сильное торможение и увеличение длины разбега. Могут создаться такие условия, что воздушное судно не д остигнет скорости отрыва и возникнет опасная ситуация.
9. Выпадающий в зимнее время снег т ребует проведения д ополнительных работ по его уборке и уплотнению на ВПП, рулежных д орожках и стоянках, где обсл уживаются воздушные суда и другие машины и механизмы.
 Как навсегда отказаться от алкоголя Плюсы полного отказа от алкоголя
Как навсегда отказаться от алкоголя Плюсы полного отказа от алкоголя Роль витаминов в лечении гриппа
Роль витаминов в лечении гриппа Традиции празднования Святого вечера накануне Рождества
Традиции празднования Святого вечера накануне Рождества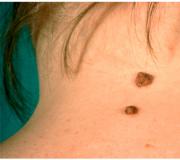 Может ли бородавка пройти сама и исчезнуть
Может ли бородавка пройти сама и исчезнуть
Влияние температуры на рост растений. Влияние температуры в доме на растения Влияние температуры на растения кратко
Негативное влияние холода зависит от диапазона понижения температур и продолжительности их воздействия. Уже неэкстремальные низкие температуры неблагоприятно сказываются на растениях, поскольку:
- тормозят основные физиологические процессы (фотосинтез, транспирацию, водообмен и т.д.),
- снижают энергетическую эффективность дыхания,
- изменяют функциональную активность мембран,
- приводят к преобладанию в обмене веществ гидролитических реакций.
Внешне повреждение холодом сопровождается потерей листьями тургора и изменением их окраски из-за разрушения хлорофилла. Основная причина повреждающего действия низкой положительной температуры на теплолюбивые растения - нарушение функциональной активности мембран из-за перехода насыщенных жирных кислот из жидкокристаллического состояния в гель. В результате, с одной стороны, повышается проницаемость мембран для ионов, а с другой - увеличивается энергия активации ферментов, связанных с мембраной. Скорость реакций, катализируемых мембранными ферментами, снижается после фазового перехода быстрее, чем скорость реакций, связанных с растворимыми энзимами. Все это приводит к неблагоприятным сдвигам в обмене веществ, резкому возрастанию количества эндогенных токсикантов, а при длительном действии низкой температуры - к гибели растения.
Установлено, что действие низких отрицательных температур находится в зависимости от состояния растений и, в частности, от оводненности тканей организма. Так, сухие семена могут выносить понижение температуры до -196°С (температура жидкого азота). Это показывает, что губительное влияние низкой температуры принципиально отлично от влияния высокой температуры, вызывающей непосредственное свертывание белков.
Основное повреждающее влияние на растительный организм оказывает льдообразование. При этом лед может образовываться как в самой клетке, так и вне клетки . При быстром понижении температуры образование льда происходит внутри клетки (в цитоплазме, вакуолях). При постепенном снижении температуры кристаллы льда образуются в первую очередь в межклетниках. Плазмалемма препятствует проникновению кристаллов льда внутрь клетки. Содержимое клетки находится в переохлажденном состоянии. В результате первоначального образования льда вне клеток водный потенциал в межклеточном пространстве становится более отрицательным по сравнению с водным потенциалом в клетке. Происходит перераспределение воды. Равновесие между содержанием воды в межклетниках и в клетке достигается благодаря:
- либо оттоку воды из клетки,
- либо образованию внутриклеточного льда.
Если скорость оттока воды из клетки соответствует скорости понижения температуры, то внутриклеточный лед не образуется. Однако гибель клетки и организма в целом может происходить в результате того, что образовавшиеся в межклетниках кристаллы льда, оттягивая воду из клетки, вызывают ее обезвоживание и одновременно оказывают на цитоплазму механическое давление, повреждающее клеточные структуры. Это вызывает ряд последствий:
- потерю тургора,
- повышение концентрации клеточного сока,
- резкое уменьшение объема клеток,
- сдвиг значений рН в неблагоприятную сторону.
Устойчивость растений к низким температурам подразделяют на холодостойкость и морозоустойчивость.
Холодостойкость растений – способность теплолюбивых растений переносить низкие положительные температуры. Защитное значение при действии низких положительных температур на теплолюбивые растения имеет ряд приспособлений. Прежде всего, это поддержание стабильности мембран и предотвращение утечки ионов . Устойчивые растения отличаются большей долей ненасыщенных жирных кислот в составе фосфолипидов мембран. Это позволяет поддерживать подвижность мембран и предохраняет от разрушений. В этой связи большую роль выполняют ферменты ацетилтрансферазы и десатуразы. Последние приводят к образованию двойных связей в насыщенных жирных кислотах.
Приспособительные реакции к низким положительным температурам проявляются в способности поддерживать метаболизм при ее снижении. Это достигается более широким температурным диапазоном работы ферментов, синтезом протекторных соединений. У устойчивых растений возрастает роль пентозофосфатного пути дыхания, эффективность работы антиоксидантной системы, синтезируются стрессовые белки. Показано, что при действии низких положительных температур индуцируется синтез низкомолекулярных белков.
Для повышения холодостойкости используется предпосевное замачивание семян. Эффективным является и использование микроэлементов (Zn, Mn, Сu, В, Мо). Так, замачивание семян в растворах борной кислоты, сульфата цинка или сульфата меди повышает холодоустойчивость растений.
Морозоустойчивость растений – способность растений переносить отрицательные температуры.
Адаптации растений к отрицательным температурам . Существуют два типа приспособлений к действию отрицательных температур:
- уход от повреждающего действия фактора (пассивная адаптация),
- повышение выживаемости (активная адаптация).
Уход от повреждающего действия низких температур достигается, прежде всего, за счет короткого онтогенеза – это уход во времени . У однолетних растений жизненный цикл заканчивается до наступления отрицательных температур. Эти растения до наступления осенних холодов успевают дать семена.
Большая часть многолетников теряет свои надземные органы и перезимовывает в виде луковиц, клубней или корневищ, хорошо защищенных от мороза слоем почвы и снега – это уход в пространстве от повреждающего действия низких температур.
Закаливание – это обратимое физиологическое приспособление к неблагоприятным воздействиям, происходящее под влиянием определенных внешних условий, относится к активной адаптации. Физиологическая природа процесса закаливания к отрицательным температурам была раскрыта благодаря работам И.И. Туманова и его школы.
В результате процесса закаливания морозоустойчивость организма резко повышается. Способностью к закаливанию обладают не все растительные организмы, она зависит от вида растения, его происхождения. Растения южного происхождения к закаливанию не способны. У растений северных широт процесс закаливания приурочен лишь к определенным этапам развития.
Закаливание растений проходит в две фазы:
Первая фаза закаливания проходит на свету при несколько пониженных плюсовых температурах (днем около 10°С, ночью около 2°С) и умеренной влажности. В эту фазу продолжается дальнейшее замедление, и даже полная остановка ростовых процессов.
Особенное значение в развитии устойчивости растений к морозу в эту фазу имеет накопление веществ-криопротекторов, выполняющих защитную функцию: сахарозы, моносахаридов, растворимых белков и др. Накапливаясь в клетках, сахара повышают концентрацию клеточного сока, снижают водный потенциал. Чем выше концентрация раствора, тем ниже его точка замерзания, поэтому накопление сахаров стабилизирует клеточные структуры, в частности хлоропласты, благодаря чему они продолжают функционировать.
Вторая фаза закаливания протекает при дальнейшем понижении температуры (около 0°С) и не требует света. В связи с этим для травянистых растений она может протекать и под снегом. В эту фазу происходит отток воды из клеток, а также перестройка структуры протопласта. Продолжается новообразование специфических, устойчивых к обезвоживанию белков. Важное значение имеет изменение межмолекулярных связей белков цитоплазмы. При обезвоживании, происходящем под влиянием льдообразования, происходит сближение белковых молекул. Связи между ними рвутся и не восстанавливаются в прежнем виде из-за слишком сильного сближения и деформации белковых молекул. В связи с этим большое значение имеет наличие сульфгидрильных и других гидрофильных группировок, которые способствуют удержанию воды и препятствуют сближению молекул белка. Перестройка цитоплазмы способствует увеличению ее проницаемости для воды. Благодаря более быстрому оттоку воды уменьшается опасность внутриклеточного льдообразования.
По отношению к температуре различают следующие типы растений:
- 1. Термофилы , мегатермные, теплолюбивые растения, температурный оптимум которых лежит в области повышенных температур.
- 2. Криофилы , микротермные, холодолюбивые растения, температурный оптимум которых лежит в области низких температур.
- 3. Мезотермные растения - промежуточная группа.
Выносливость растений к экстремальным температурам характеризует их жаростойкость и морозостойкость. На действие температуры как фактора, наземные растения выработали ряд адаптаций.
Так, от перегрева растение защищает:
- 1. Транспирация (при испарении 1 г воды при 20° требуется 500 ккал)
- 2. Блестящая поверхность, густое опушение, вертикальное расположение узких листовых пластинок (типчак, ковыль), общая редукция листовой поверхности - то есть все те приспособления, которые служат для ослабления влияния солнечной радиации.
- 3. Пробка на коре, воздушные полости на корневой шейке - приспособления, свойственные пустынным растениям.
- 4. Своеобразной адаптацией является занятие растениями определенных экологических ниш, защищенных от перегрева.
- 5. Переживание наиболее жарких месяцев в состоянии анабиоза или в виде семян и подземных органов.
Специальных адаптации к действию холода у растений нет, но от всего комплекса неблагоприятных факторов, с ним связанных (сильные ветры, возможность иссушения) растение защищают такие морфологические особенности как опушение почечных чешуй, засмоление почек, утолщенный пробковый слой, толстая кутикула. Своеобразное приспособление к холоду наблюдается в высокогорьях Африки у розеточных деревьев лобелии во время ночного холода розетки листьев закрываются.
Защите от холода способствуют также:
- 1. Малые размеры, карликовость, или нанизм . Например, у карликовой березы и ивы - Веtula nana, Salix роlaris.
- 2. Стелющиеся формы - стланцы .
- 3. Переживание наиболее жарких месяцев в состоянии анабиоза или в виде семян, или подземных органов.
- 4. Особая жизненная форма растений-подушек (у вереска) способна поддерживать температуру в гуще ветвей на 13°С выше окружающей.
- 5. Развитие контрактильных - сократительных корней. Осенью такие корни высыхают, сокращаются и вдавливают зимующие почки вглубь почвы, что препятствует выталкивающей силе вечной мерзлоты).
Для растений умеренных областей более характерны физиологические способы защиты от холода.
- 1. Снижение точки замерзания клеточного сока (больше растворимых сахаров, увеличение доли коллоидно-связанной воды). В целом растения в этом плане хуже приспособлены чем насекомые.
- 2. Снижение температурных оптимумов физиологических процессов. У арктических лишайников, например, фотосинтез оптимален при 5° и возможен при -10°
- 3. Подснежный рост в предвесенний период у пролесков, тюльпанов и других эфемероидов.
- 4. Анабиоз - крайняя мера защиты растений - состояние покоя, во время которого растение способно вынести до -200°С. В состоянии зимнего покоя различают фазу глубокого или органического покоя, когда срезанные ветки не распускаются в тепле и фазу вынужденного покоя в конце зимы. Сигналом к наступлению покоя служит сокращение дня.
Министерство образования Республики Беларусь
Учреждение образования БГПУ им. М.Танка
по физиологии растений
на тему: «Влияние перегрева растений на их функциональные особенности»
Влияние высоких температур на растения
Для большинства растений наиболее благоприятными для жизни являются температуры +15…+30 о С. При температуре +35…+40 о С большинство растений повреждаются.
Действие высоких температур влечет за собой целый ряд опасностей для растений: сильное обезвоживание и иссушение, ожоги, разрушение хлорофилла, необратимые расстройства дыхания и других физиологических процессов, прекращение синтеза белков и усиление их распада, накопление ядовитых веществ, в частности аммиака. При очень высоких температурах резко повышается проницаемость мембран, а затем наступает тепловая денатурация белков, коагуляция цитоплазмы и отмирание клеток. Перегрев почвы приводит к повреждению и отмиранию поверхностно расположенных корней, к ожогам корневой шейки.
Первичные изменения клеточных структур происходят на уровне мембран в результате активации образования кислородных радикалов и последующего перекисного окисления липидов, нарушения антиоксидантной системы – активности супероксиддисмутазы, глутатионредуктазы и других ферментов. Это вызывает разрушение белково-липидных комплексов плазмалеммы и других клеточных мембран, приводит к потере осмотических свойств клетки. В результате наблюдаются дезорганизация многих функций клеток, снижение скорости различных физиологических процессов. Так, при температуре 20 о С все клетки проходят процесс митотического деления, при 38 о С митоз отмечается в каждой седьмой клетке, а повышение температуры до 42 о С снижает число делящихся клеток в 500 раз.
При максимальных температурах расход органических веществ на дыхание превышает его синтез, растение беднеет углеводами, а затем начинает голодать. Особенно резко это выражено у растений более умеренного климата (пшеница, картофель, многие огородные культуры). При общем ослаблении повышается их восприимчивость к грибковым и вирусным инфекциям.
Даже кратковременное стрессирующее действие высокой температуры вызывает перестройку гормональной системы растений. На примере проростков пшеницы и гороха установлено, что тепловой шок индуцирует целый каскад многоступенчатых изменений гормональной системы, который запускается выбросом ИУК из пула ее конъюгатов, выполняющего роль стрессового сигнала и инициирующего синтез этилена. Результат синтеза этилена – последующее снижение уровня ИУК и увеличение АБК. Эти гормональные перестройки, очевидно, индуцируют синтез ферментов антиоксидантной защиты и белков теплового шока, вызывают снижение темпов роста и как следствие – повышается устойчивость растения к действию высоких температур.
Существует определенная связь между условиями местообитания растений и жароустойчивостью. Чем суше местообитание, тем выше температурный максимум, тем больше жароустойчивость растений.
К воздействию высоких температур растения могут подготовиться за несколько часов. Так, в жаркие дни устойчивость растений к высоким температурам после полудня выше, чем утром. Обычно эта устойчивость временная, она не закрепляется и довольно быстро исчезает, если становится прохладно. Обратимость теплового воздействия может составлять от нескольких часов до 20 дней.
Жароустойчивость связана также со стадией развития растений: молодые, активно растущие ткани менее устойчивы, чем старые. Особенно опасны высокие температуры в период цветения. Практически все генеративные клетки в данных условиях претерпевают структурные изменения, теряют активность и способность к делению, наблюдается деформация пыльцевых зерен, слабое развитие зародышевого мешка и появление стерильных цветков.
Отличаются по жароустойчивости и органы растений. Лучше переносят повышенную температуру обезвоженные органы: семена до 120 о С, пыльца до 70 о С, споры в течении нескольких минут выдерживают нагревание до 180 о С.
Из тканей наиболее устойчивы камбиальные. Так, камбиальный слой в стволах переносит летом температуру до +51 о С.
Приспособления растений к высоким температурам
перегрев растение температура жароустойчивость
Жароустойчивость – это способность теплолюбивых растений длительно, а умеренно теплолюбивых кратковременно переносить действие высоких температур, перегрев.
В защитных приспособлениях растений к высоким температурам использованы разные пути адаптации.
Морфологические черты : в основном те же, что служат растению для ослабления прихода солнечной радиации к тканям надземных частей и обеспечивают возможность уменьшения потерь воды.
1. усиленная транспирация. Необходимо отметить, что у интенсивно транспирирующих видов охлаждение листьев достигает 15 о С. Это крайний пример, но и снижение на 3-4 о С может предохранить от губительного перегрева.
2. стабилизация метаболических процессов (более жесткая структура мембран, высокая вязкость цитоплазмы, низкое содержание воды в клетке и т.д.). Под действие температуры, прежде всего, изменяется содержание липидов мембран. Так, теплоустойчивые сорта люцерны при +30 о С содержали большое количество сульфо- и фосфолипидов, чем при +15 о С. Кроме того, повышение температуры оказывает влияние и на жирнокислотный состав липидов: увеличивается содержание насыщенных жирных кислот, более тугоплавких.
3. высокая интенсивность фотосинтеза и дыхания.
4. высокое содержание защитных веществ (слизи, органических кислот и др.). Аммиак, образовавшийся при распаде белка, вызывает отравление растительных клеток и их гибель. Под влиянием высоких температур у жароустойчивых растений снижается дыхательный коэффициент и накапливаются органические кислоты, которые обезвреживают аммиак, образуя с ним аммонийные соли. Кроме этого, аммиак связывается с аминокислотами с образованием амидов и с аланином, синтез которого при +30-40 о С резко возрастает.
5. сдвиги температурного оптимума активности важнейших ферментов.
6. синтез термостойких белков (БТШ)
БТШ обнаруживаются в клетках через 10-15 мин после повышения температуры, а через 0,5-3,5 ч наблюдается их максимальное содержание. Локализуются данные белки в ядре, цитозоле, клеточных органеллах и функционируют в клетках в виде высокомолекулярных комплексов. Очевидно, большинство низкомолекулярных белков теплового шока выполняют функции шаперонов, т.е. защищают полипептиды от денатурации в период стресса и восстанавливают поврежденные белки. Действие белков теплового шока приурочено к начальному периоду ответа растений на повышение температуры, т.е. БТШ защищают клетки лишь в течение очень ограниченного времени. Синтез БТШ имеет кратковременный характер, т.к. их длительный синтез невозможен из-за крайне высокой потребности в энергии. Однако БТШ, предотвращая быструю гибель растения создают тем самым условия для формирования более совершенных долговременных механизмов адаптации.
Рассматривая приспособления растений к действию высоких температур, необходимо отметить своеобразное физиологическое приспособление к температуре среды, превышающей адаптивные возможности растений, - переход в состояние анабиоза. Из этого состояния живые существа могут возвратиться к нормальной активности только в том случае, если не была нарушена структура макромолекул в их клетках.
Существует еще один способ адаптации растений к чрезмерно высоким температурам – смещение вегетации на сезон с более благоприятными температурными условиями. Это сезонная адаптация, связанная с перестройкой всего годичного цикла развития, обеспечивает растениям надежную защиту от жары даже в районах самых жарких пустынь.
Экологические группы растений по жароустойчивости
Нежаростойкие – мезофитные и водные растения. Борются с перегревом с помощью вертикального расположения листьев, складывания и свертывания листовых пластинок, увеличением интенсивности транспирации. Более жаростойкие мезофиты отличаются повышенной вязкостью цитоплазмы и концентрацией клеточного сока, усиленным синтезом жаростойких белков-ферментов.
Жаровыносливые – растения пустынь и сухих мест обитания. Они характеризуются специфическими морфолого-анатомическими особенностями строения отдельных органов, имеют пониженный уровень метаболических процессов, отличаются повышенной вязкостью цитоплазмы, высоким содержанием связанной воды в клетке и т.д.
Некоторые растения в условиях жаркого климата способны выделять соли, из которых на стволах и листьях образуются кристаллы, преломляющие и отражающие падающие лучи солнца.
Жаростойкие – термофильные сине-зеленые водоросли и бактерии горячих минеральных источников и кратеров вулканов. Жароустойчивость определяется высоким уровнем метаболизма, повышенным содержанием РНК в клетках, устойчивостью белка цитоплазмы и тепловой денатурации, синтезом более жароустойчивых белков-ферментов, высокой вязкостью цитоплазмы, повышенным содержанием осмотически активных веществ.
1. Жукова И.И. Адаптация растений к условиям окружающей среды. Могилев, 2008.
2. Физиология и биохимия сельскохозяйственных растений/ Третьяков Н.Н. и др. – М.: Колос, 2000.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Министерство образования Российской Федерации
Государственное образовательное учреждение
высшего профессионального образования
ИРКУТСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Влияние температуры на растения
доцент, к.б.н. Машанова О.Я.
учебная группа 6141
Приспособленность онтогенеза растений к условиям среды является результатом их эволюционного развития (изменчивости, наследственности, отбора). На протяжении филогенеза каждого вида растений в процессе эволюции выработались определенные потребности индивидуума к условиям существования и приспособленность к занимаемой им экологической нише. Влаголюбие и теневыносливость, жароустойчивость, холодоустойчивость и другие экологические особенности конкретных видов растений сформировались в ходе эволюции в результате длительного действия соответствующих условий. Так, теплолюбивые растения и растения короткого дня характерны для южных широт, менее требовательные к теплу и растения длинного дня - для северных.
В природе в одном географическом регионе каждый вид растений занимает экологическую нишу, соответствующую его биологическим особенностям: влаголюбивые - ближе к водоемам, теневыносливые - под пологом леса и т. д. Наследственность растений формируется под влиянием определенных условий внешней среды. Большое значение имеют и внешние условия онтогенеза растений.
В большинстве случаев растения и посевы (посадки) сельскохозяйственных культур, испытывая действие тех или иных неблагоприятных факторов, проявляют устойчивость к ним как результат приспособления к условиям существования, сложившимся исторически.
1. Температура как биологический фактор
Растения - пойкилотермные организмы, т.е. их собственная температура уравнивается с температурой окружающей их среды. Однако это соответствие неполное. Конечно, тепло, выделяемое при дыхании и используемое при синтезах, вряд ли играет какую-либо экологическую роль, но все же температура надземных частей растения может значительно отличаться от температуры воздуха в результате энергообмена с окружающей средой. Благодаря этому, например, растения Арктики и высокогорий, которые заселяют места, защищенные от ветра, или растут вплотную к почве, имеют более благоприятный тепловой режим и могут достаточно активно поддерживать обмен веществ и рост, несмотря на постоянно низкие температуры воздуха. Не только отдельные растения и их части, но и целые фитоценозы обнаруживают иногда характерные отклонения от температуры воздуха. В один жаркий летний день в Центральной Европе температура на поверхности крон в лесах была на 4°С, а лугов - на 6 °С выше температуры воздуха и на 8 °С (лес) или 6 °С (луг) ниже, чем температура поверхности почвы, лишенной растительности.
Чтобы охарактеризовать тепловые условия местообитания растений, необходимо знать закономерности распределения тепла в пространстве и его динамику во времени как в отношении общеклиматических характеристик, так и конкретных условий произрастания растений.
Общее представление об обеспеченности того или иного района теплом дают такие общеклиматические показатели, как среднегодовая температура для данной местности, абсолютный максимум и абсолютный минимум (т.е. наиболее высокая и наиболее низкая температура, зарегистрированная в этом районе), средняя температура самого теплого месяца (на большей части северного полушария это июль, южного полушария - январь, на островах и в прибрежных районах - август и февраль); средняя температура самого холодного месяца (в континентальных областях северного полушария - январь, южного - июль, в прибрежных районах - февраль и август).
Для характеристики тепловых условий жизни растений важно знать не только общее количество тепла, но и его распределение во времени, от которого зависят возможности вегетационного периода. Годовую динамику тепла хорошо отражает ход среднемесячных (или среднесуточных) температур, неодинаковый на разных широтах и при разных типах климата, а также динамика максимальных и минимальных температур. Границы вегетационного сезона определяются продолжительностью безморозного периода, частотой и степенью вероятности весенних и осенних заморозков. Естественно, порог вегетации не может быть одинаковым для растений с разным отношением к теплу; для холодостойких культурных видов условно принимают 5°С, для большинства культур умеренной зоны 10°С, для теплолюбивых 15°С. Считают, что для естественной растительности умеренных широт пороговая температура начала весенних явлений составляет 5°С.
В общих чертах скорость сезонного развития пропорциональна накопленной сумме температур (стоит, например, сравнить медленное развитие растений в холодную и затяжную весну или «взрывное» начало весны при сильной волне тепла). От этой общей закономерности есть ряд отступлений: так, например, слишком высокие суммы температур уже не ускоряют, а тормозят развитие.
2. Температура растений
Наряду с тепловыми характеристиками окружающей среды необходимо знать температуру самих растений и ее изменения, поскольку именно она представляет истинный температурный фон для физиологических процессов. Температуру растений измеряют с помощью электротермометров, имеющих миниатюрные полупроводниковые датчики. Чтобы датчик не повлиял на температуру измеряемого органа, необходимо, чтобы его масса была во много раз меньше массы органа. Датчик должен быть также малоинерционным и быстро реагировать на изменения температуры. Иногда для этой цели используют термопары. Датчики или прикладывают к поверхности растения, или «вживляют» в стебли, листья, под кору (например, для измерения температуры камбия). Одновременно обязательно измеряют температуру окружающего воздуха (затенив датчик).
Температура растений весьма непостоянна. Из-за турбулентных потоков и непрерывных изменений температуры воздуха, непосредственно окружающего лист, действия ветра и т. д. температура растения варьирует с размахом в несколько десятых долей или даже целых градусов и с частотой в несколько секунд. Поэтому под «температурой растений» следует понимать более или менее обобщенную и в достаточной мере условную величину, характеризующую общий уровень нагрева. Растения как пойкилотермные организмы не имеют собственной стабильной температуры тела. Их температура определяется тепловым балансом, т. е. соотношением поглощения и отдачи энергии. Эти величины зависят от многих свойств как окружающей среды (размеры прихода радиации, температура окружающего воздуха и его движение), так и самих растений (окраска и другие оптические свойства растения, величина и расположение листьев и т. д.). Первостепенную роль играет охлаждающее действие транспирации, которое препятствует очень сильным перегревам в жарких местообитаниях. Это легко показать в опытах с пустынными растениями: стоит лишь смазать вазелином ту поверхность листа, на которой расположены устьица, и лист на глазах гибнет от перегрева и ожогов.
В результате действия всех указанных причин температура растений обычно отличается (иногда довольно значительно) от температуры окружающего воздуха. При этом возможны три ситуации:
· температура растения выше температуры окружающего воздуха («супратемпературные» растения, по терминологии О. Ланге),
· ниже ее («субтемпературные»),
· равна или очень близка к ней.
Первая ситуация встречается довольно часто в самых разнообразных условиях. Значительное превышение температуры растения над температурой воздуха обычно наблюдается у массивных органов растений, особенно в жарких местообитаниях и при слабой транспирации. Сильно нагреваются крупные мясистые стебли кактусов, утолщенные листья молочаев, очитков, молодила, у которых испарение воды очень незначительное. Так, при температуре воздуха 40- 45°С пустынные кактусы нагреваются до 55- 60°С; в умеренных широтах в летние дни сочные листья растений из родов Sempervivum и Sedum нередко имеют температуру 45°С, а внутри розеток молодила - до 50°С. Таким образом, превышение температуры растения над температурой воздуха может достигать 20°С.
Сильно нагреваются солнцем различные мясистые плоды: например, спелые томаты и арбузы на 10- 15°С теплее воздуха; температура красных плодов в зрелых початках аронника - Arum maculatum доходит до 50°С. Довольно заметно бывает повышение температуры внутри цветка с более или менее закрытым околоцветником, сохраняющим от рассеивания тепло, которое выделяется при дыхании. Иногда это явление может иметь существенное адаптивное значение, например, для цветков лесных эфемероидов (пролески, хохлатки и др.), ранней весной, когда температура воздуха едва превышает 0°С.
Своеобразен и температурный режим таких массивных образований, как древесные стволы. У одиночно стоящих деревьев, а также в лиственных лесах в «безлистную» фазу (весной и осенью) поверхность стволов сильно нагревается в дневные часы, причем в наибольшей степени с южной стороны; температура камбия здесь может быть на 10- 20°С выше, чем на северной стороне, где она имеет температуру окружающего воздуха. В жаркие дни температура темных стволов ели повышается до 50- 55°С, что может принести к ожогам камбия. Показания тонких термопар, вживленных под кору, позволили установить, что стволы древесных пород защищены по-разному: у березы температура камбия быстрее меняется в соответствии с колебаниями температуры наружного воздуха, в то время как у сосны она более постоянна благодаря лучшим теплозащитным свойствам коры. Нагревание стволов деревьев и безлистном весеннем лесу существенно влияет на микроклимат лесного сообщества, поскольку стволы - хорошие аккумуляторы тепла.
Превышение температуры растений над температурой воздуха встречается не только в сильно прогреваемых, но и в более холодных местообитаниях. Этому способствует темная окраска или иные оптические свойства растений, увеличивающие поглощение солнечной радиации, а также анатомо-морфологические особенности, способствующие снижению транспирации. Довольно заметно могут нагреваться арктические растения: один пример - карликовая ива - Salix arctica на Аляске, у которой днем листья теплее воздуха на 2- 11°С и даже в ночные часы полярного «круглосуточного дня» - на 1- 3°С. Еще один интересный пример нагревания под снегом: в летнее время в Антарктиде температура лишайников бывает выше 0°С даже под слоем снега более 30 см. Очевидно, в столь суровых условиях естественный отбор сохранил формы с наиболее темной окраской, у которых благодаря такому нагреванию возможен положительный баланс углекислотного газообмена.
Довольно значительно могут нагреваться солнечными лучами иглы хвойных древесных пород зимой: даже при отрицательных температурах возможно превышение над температурой воздуха на 9- 12°С, что создает благоприятные возможности для зимнего фотосинтеза. Экспериментально было показано, что если для растений создать сильный поток радиации, то даже при низкой температуре порядка - 5, - 6°С листья могут нагреться до 17- 19°С, т. е. фотосинтезировать при вполне «летних» температурах.
Снижение температуры растений по сравнению с окружающим воздухом чаще всего отмечается в сильно освещенных и прогреваемых местообитаниях (степях, пустынях), где листовая поверхность растений сильно редуцирована, а усиленная транспирация способствует удалению избытка тепла и предотвращает перегрев. У интенсивно транспирирующих видов охлаждение листьев (разность с температурой воздуха) достигает 15°С. Это крайний пример, но и снижение на 3- 4°С может предохранить от губительного перегрева.
В самых общих чертах можно сказать, что в жарких местообитаниях температура надземных частей растений ниже, а в холодных- выше температуры воздуха. Эта закономерность прослеживается и на одних и тех же видах: так, в холодном поясе гор Северной Америки, на высотах 3000- 3500 м, растения теплее, а в низкогорном - холоднее воздуха.
Совпадение температуры растений с температурой окружающего воздуха встречается гораздо реже в условиях, исключающих сильный приток радиации и интенсивную транспирацию, например у травянистых растений под пологом тенистых лесов (но не на солнечных бликах), а на открытых местообитаниях - в пасмурную погоду или при дожде.
Различают разные биологические типы растений по отношению к температуре. У растений термофильных, или мегатермных (теплолюбивых), оптимум лежит в области повышенных температур. Они обитают в областях тропического и субтропического климата, а в умеренных поясах - в сильнопрогреваемых местообитаниях. Для криофильных,или микротермных (холодолюбивых), растений оптимальны низкие температуры. К ним принадлежат виды, живущие в полярных и высокогорных областях или занимающие холодные экологические ниши. Иногда выделяют промежуточную группу мезотермных растений.
3. Действие температурного стресса
Жара и мороз вредят жизненным функциям и ограничивают распространение вида в зависимости от их интенсивности, продолжительности и периодичности, но прежде всего от состояния активности и степени закалки растений. Стресс-- это всегда необычная нагрузка, которая не обязательно должна быть опасной для жизни, но которая непременно вызывает в организме «реакцию тревоги», если только он не находится в выраженном состоянии оцепенения. Покоящиеся стадии, такие, как сухие споры, а также пойкилогидрические растения в высохшем состоянии, нечувствительны, так что они могут пережить без повреждения любую отмеченную на Земле температуру.
Протоплазма вначале отвечает на стресс резким усилением метаболизма. Повышение интенсивности дыхания, которое наблюдается в качестве стрессовой реакции отражает попытку исправить уже появившиеся дефекты и создать ультраструктурные предпосылки для приспособления к новой ситуации. Стрессовая реакция - это борьба механизмов адаптации, с деструктивными процессами в протоплазме, ведущими к ее гибели.
Гибель клеток от перегрева и холода
Если температура переходит критическую точку, клеточные структуры и функции могут повреждаться так внезапно, что протоплазма тотчас же отмирает. В природе такое внезапное разрушение нередко происходит при эпизодических морозах, например при поздних заморозках весной. Но повреждения могут возникать и постепенно; отдельные жизненные функции выводятся из равновесия и угнетаются, пока, наконец, клетка не отомрет в результате прекращения жизненноважных процессов.
3.1 Картина повреждения
Различные жизненные процессы неодинаково чувствительны к температуре. Сначала прекращается движение протоплазмы, интенсивность которого непосредственно зависит от энергоснабжения за счет процессов дыхания и от наличия высокоэнергетических фосфатов. Затем снижаются фотосинтез и дыхание. Для фотосинтеза особенно опасна жара, дыхание же наиболее чувствительно к холоду. У поврежденных холодом или жарой растений после возвращения в умеренные условия уровень дыхания сильно колеблется и часто бывает ненормально повышен. Повреждение хлоропластов ведет к длительному или необратимому угнетению фотосинтеза. В конечной стадии утрачивается полупроницаемость биомембран, разрушаются клеточные компартменты, особенно тилакоиды пластид, и клеточный сок выходит в межклетники..
3.2 Причины гибели при перегреве
Высокая температура быстро приводит к гибели из-за повреждения мембран и прежде всего в результате инактивации и денатурации белков. Даже если из строя выходят только немногие, особо термолабильные ферменты, это ведет к расстройству обмена нуклеиновых кислот и белков и в конце концов - тоже к гибели клеток. Растворимые азотистые соединения накапливаются при этом в таких больших концентрациях, что они диффундируют из клеток и теряются; кроме того, образуются ядовитые продукты распада, которые не могут больше обезвреживаться в ходе обмена веществ.
3.3 Гибель от охлаждения и от мороза
растение температура перегрев мороз
При повреждении протоплазмы холодом следует различать, вызвано ли оно самой по себе низкой температурой или же замерзанием. Некоторые растения тропического происхождения повреждаются уже при снижении температуры до нескольких градусов выше нуля. Подобно гибели от перегрева, гибель от охлаждения тоже бывает прежде всего связана с дезорганизацией обмена нуклеиновых кислот и белков, но здесь играют роль также нарушения, проницаемости и прекращение тока ассимилятов.
Растения, которым охлаждение до температур выше нуля не причиняет вреда, повреждаются только при температурах ниже нуля, т. е. в результате образования льда в тканях. Богатые водой, незакаленные протопласты могут легко замерзать; при этом внутри клетки мгновенно образуются ледяные кристаллики, и клетка погибает. Чаще всего лед образуется не в протопластах, а в межклетниках и клеточных стенках. Такое образование льда называют внеклеточным. Выкристаллизовавшийся лед действует как сухой воздух, так как упругость пара надо льдом ниже, чем над переохлажденным раствором. В результате от протопластов отнимается вода, они сильно сжимаются (на 2/з своего объема) и концентрация растворенных веществ в них возрастает. Перемещение воды и замерзание продолжаются до тех пор, пока в протоплазме не установится равновесие сосущих сил между льдом и водой. Положение равновесия зависит от температуры; при температуре - 5°С равновесие наступает примерно при; 60 бар, а при - 10°С - уже при 120 -бар. Таким образом, низкие температуры действуют на протоплазму так же, как высыхание. Морозоустойчивость клетки более высока, если вода прочно связана со структурами протоплазмы и осмотически связана. При обезвоживании цитоплазмы (безразлично, в результате засухи или замерзания) инактивируются ферментные системы, ассоциированные с мембранами, - системы, участвующие главным образом в синтезе АТФ и в процессах фосфорилирования (Хебер, Сантариус, 1979). Инактивацию, вызывают чрезмерные и потому токсичные концентрации ионов. солей и органических кислот в незамерзшем остаточном растворе. Напротив, сахара, производные сахаров, определенные аминокислоты и белки защищают биомембраны и ферменты от вредных веществ (Максимов, Туманов, Красавцев, 1952). Наряду с этим есть указания на то, что при замерзании белки денатурируются, что также ведет к повреждению мембран (Левитт 1980).
Термоустойчивость - это способность организма переносить большую жару или холод без необратимого повреждения. Термоустойчивость растения складывается из способности протоплазмы переносить экстремальные температуры (толерантность по Дж. Левитту) и из эффективности мер, которые замедляют или предотвращают развитие повреждений (избегание).
Меры, позволяющие избегать повреждения
Возможные способы защиты клеток от температурного повреждения немногочисленны и не очень эффективны. Изоляция от перегрева и охлаждения может дать лишь кратковременную защиту. Так, например, в густых кронах деревьев или у подушечных растений почки листья и цветки, находящиеся в глубине и ближе к земле, меньше подвергаются опасности замерзнуть в результате отдачи тепла путем излучения, чем наружные части растения. Виды хвойных с особенно толстой коркой лучше выдерживают пожары в подлеске. Общее же значение имеют главным образом две защитные меры: замедление образования льда в тканях и (при жаре) охлаждение путем отражения падающих лучей и с помощью транспирации.
3.5 Устойчивость протоплазмы
Длительное и регулярно повторяющееся воздействие крайних температур растения могут выдерживать только в том случае, если сама протоплазма жаро- или морозоустойчива. Эта особенность обусловлена генетически и поэтому у разных видов и даже сортов выражена в разной степени. Однако это не такое свойство, которое присуще растению постоянно и всегда в одинаковой мере. Проростки, весенние побеги древесных растений в период их интенсивного растяжения, культуры микроорганизмов в фазе экспоненциального роста вряд ли способны закаливаться и, поэтому чрезвычайно чувствительны к температуре.
Устойчивость к образованию льда и закаливание к действию мороза
В областях с сезонным климатом наземные растения приобретают осенью «льдоустойчивость», т. Е. способность переносить образование льда в тканях. Весной, с распусканием почек они снова утрачивают эту способность, и теперь замерзание приводит к их вымерзанию. Таким образом, холодостойкость многолетних растений вне тропиков регулярно колеблется в течение года между минимальной величиной в период вегетации и максимумом в зимнее время. Льдоустойчивость формируется постепенно осенью. Первая предпосылка для этого - переход растения в состояние готовности к закаливанию, наступающее только тогда, когда заканчивается рост. Если готовность к закаливанию достигнута, то процесс закаливания может начинаться. Этот процесс состоит из нескольких фаз, каждая из которых подготавливает переход к следующей. Закаливание к морозу, у озимых злаков и плодовых; деревьев (эти растения изучались наиболее основательно) начинается многодневным (до нескольких недель) воздействием температур чуть выше нуля. На этой фазе, предшествующей закаливанию, в протоплазме накапливаются сахара и другие защитные вещества, клетки становятся беднее водой, а центральная вакуоль распадается на множество мелких вакуолей. Благодаря этому протоплазма оказывается подготовленной к следующей фазе, проходящей при регулярных слабых морозах от - 3 до- 5°С. При этом ультраструктуры и ферменты протоплазмы перестраиваются таким образом, что клетки переносят обезвоживание, связанное с образованием льда. Только после этого растения могут, не подвергаясь, опасности, вступать в заключительную фазу процесса; закаливания, которая при непрерывном морозе по меньшей мере от - 10 до-15 °С делает протоплазму в высшей степени морозоустойчивой.
Зоны эффективных температур для разных видов различны. Готовые к закаливанию сеянцы березы, которые до начала процесса закаливания вымерзали бы при температуре от - 15-до - 20°С, переносят после окончания первой фазы закаливания; уже - 35 °С, а при полной закалке они выдерживают даже охлаждение до - 195°С. Таким образом, холод сам по себе стимулирует процесс закаливания. Если мороз ослабевает, то протоплазма опять переходит в первую фазу закаливания, однако устойчивость, снова может быть поднята холодными периодами до наивысшего уровня, пока растения остаются в состоянии покоя.
В зимний период на сезонный ход морозостойкости накладываются кратковременные (индуцированные), адаптации, благодаря которым уровень устойчивости быстро приспосабливается к изменениям погоды. Холод больше всего способствует закаливанию в начале зимы. В это время устойчивость может за несколько дней подняться до наивысшего уровня. Оттепель, особенно в конце зимы, вызывает быстрое снижение устойчивости растений, но в середине зимы после выдерживания в течение.нескольких дней при температуре от +10 до +20 °С растения в значительной мере теряют закалку. Способность изменять морозоустойчивость под влиянием холода и тепла, т. Е. диапазон индуцируемых адаптации устойчивости, является конституционным признаком отдельных видов растений.
После окончания зимнего покоя способность к закаливанию и вместе с тем высокая степень закалки быстро утрачиваются. Весной существует тесная связь между активированием распускания почек и ходом изменения устойчивости
Формы приспособлений у растений бесконечно разнообразны. Весь растительный мир со времени своего появления совершенствуется по пути целесообразных приспособлений к условиям обитания.
Растения - пойкилотермные организмы. Повреждения начинаются на молекулярном уровне с нарушений функций белков и нуклеиновых кислот. Температура - это фактор, серьезно влияющий на морфологию и физиологию растений, требующий изменений в самом растении, которые могли бы приспособить его. Адаптации растений к разным температурным условиям даже в пределах одного вида различны.
При высоких температурах выявлены такие адаптации, как густое опушение листьев, блестящая поверхность, уменьшение поверхности, поглощающей радиацию, изменение положения по отношению к источнику тепла, усиление транспирации, высокое содержание защитных веществ, сдвиг температурного оптимума активности важнейших ферментов, переход в состояние анабиоза, занятие микрониш, защищенных от инсоляции и перегрева, сдвиг вегетации на сезон с более благоприятными тепловыми условиями.
Адаптации к холоду таковы: опушение почечных чешуй, толстая кутикула, утолщение пробкового слоя, опушение листьев, закрывание розеточных листьев ночью, развитие карликовости, развития стелящихся форм, подушковая форма роста, развитие контрактильных корней, повышение концентрации клеточного сока, повышение доли коллоидно-связанной воды, анабиоз
По различной термоустойчивости выделяются виды: нехолодостойкие, неморозоустойчивые, льдоустойчивые, нежаростойкие, жаровыносливые зукариоты, жаровыносливые прокариоты.
Список использованной литературы
1. Александров В.Я. Клетки, макромолекулы и температура. Л.: Наука, 1975г. 328 с
2. Вознесенский В. Л., Рейнус Р. М. Температура ассимилирующих органов пустынных растений // Бот. журн., 1977; т. 62. N 6
3. Горышина Т. К. Ранневесенние эфемероиды лесостепных дубрав. Л., Изд-во Ленингр. ун-та. 1969
4. Горышина Т.Н. Экология растений уч. Пособие для ВУЗов, Москва, В.
5. Культиасов И.М. Экология растений М.: Изд-во московского ун-та, 1982 33-89с.
6. Лархер В. Экология растений М.: Мир 1978 г. 283-324c.
7. Максимов Н. А. Избранные работы по засухоустойчивости и зимостойкости растений М.:Изд-во АН-СССР.-1952 т. 1-2
8. Полевой В.В. Физиология растений 1978г. 414-424с.
9. Селянинов Г. Т. К методике сельскохозяйственной климатологии. Труды по с.-х. метеорологии, 1930, т. 22
10. Тихомиров Б. А. Очерки по биологии растений Арктики. Л., Изд-во АН СССР, 1963
11. Туманов И. И. Причины гибели растений в холодное время года и меры её предупреждения. М., Знание, 1955
Размещено на Allbest.ru
Подобные документы
Температура как экологический фактор. Температура растений. Действие температурного стресса. Картина повреждения. Причины гибели при перегреве. Гибель от охлаждения и от мороза. Устойчивость протоплазмы. Растения и высокая температура.
курсовая работа , добавлен 31.07.2007
Влияние перегрева растений на их функциональные особенности, виды опасностей. Связь между условиями местообитания растений и жароустойчивостью. Приспособления и адаптация растений к высоким температурам. Экологические группы растений по жароустойчивости.
реферат , добавлен 23.04.2011
Почему прорастание семян у разных растений происходит при разных температурах. Какое значение имеет промораживание семян растений. Что задерживает тепло в атмосфере. Продолжительность вегетационного периода. Определение температуры тела растения.
презентация , добавлен 11.04.2013
Краткие характеристики климатических условий на цветение раннецветущих растений. Суточное колебание температуры воздуха. Влияние время схода снега на сезонное развитие растений. Характеристика раннецветущих травянистых растений, кустарников и деревьев.
курсовая работа , добавлен 01.06.2014
Жизненный цикл древесных растений. Выражение приспособленности к условиям среды. Фенологическое развитие древесных растений. Программа фенологических наблюдений. Растения на ювенильном этапе онтогенеза, на виргинилъном и последующих этапах онтогенеза.
реферат , добавлен 24.02.2009
Влияние температуры на особенности прорастания и всхожести семян эфемеров в лабораторных и полевых условиях. Определение минимальной, оптимальной и максимальной температуры прорастания семян эфемерных растений Донбасса, их таксономический анализ.
магистерская работа , добавлен 19.11.2015
Причины приспособленности организмов к среде обитания. Географическое (аллопатрическое) видообразование. Действие мутационного процесса на популяцию в природе. Биологический прогресс и регресс. Ароморфоз как направление эволюции. Примеры идиоадаптаций.
презентация , добавлен 21.01.2011
Растения-индикаторы - растения, для которых характерна резко выраженная адаптация к определённым условиям окружающей среды. Реакции живых организмов на будущие изменения погодных условий. Примеры использования индикационных свойств растений и животных.
презентация , добавлен 30.11.2011
Исследование строения и основных свойств экосистем. Изучение экологических связей в естественных и искусственных экосистемах. Анализ взаимоотношений в системе "организм-среда". Пищевая цепь растений. Приспособленность растений к условиям среды обитания.
практическая работа , добавлен 23.10.2014
Определение понятий "засуха" и "засухоустойчивость". Рассмотрение реакции растений на засуху. Изучение типов растений по отношению к водному режиму: ксерофитов, гигрофитов и мезофитов. Описание механизма приспособления растений к условиям внешней среды.
Жизнь и развитие комнатных растений зависит от многих факторов и основным из них является температура. Влияние температуры на растения может быть как положительным так и крайне отрицательным. Конечно все зависит от вида растения и его предпочтений в условиях дикой природы, но некоторые виды утрачивают изначальные привычки и полностью адаптируются к условиям квартир.
Каждый тип растений нуждается в разном количестве тепла, некоторые их них могут стойко переносить отклонения от приемлемых температурных режимов, а другие страдают и тормозятся в развитии.
Важным фактором является не только количество тепла получаемого растением, но и продолжительность теплового воздействия. На разных этапах жизни растения количество необходимого тепла не редко варьируется, так на стадии активного роста большинству растений необходима теплая атмосфера, но когда растение переходит в период покоя количество получаемого тепла рекомендуется снизить.
Комфортная температура для каждого растения определяется исходя из значений максимальной и минимальной температуры при которой растение нормально развивается или комфортно себя ощущает на разных этапах жизни. Падение температуры ниже допустимых значений, как правило, приводит к затуханию всех процессов, торможению развития и ослаблению процесса фотосинтеза. Повышение напротив активизирует и ускоряет эти процессы.
В холодное время года влияние температуры на растения немного отличается. Растения будет комфортно при более низких температурах, это обусловлено тем, что большинство растений в этот период переходят в фазу покоя. В это время процесс роста замедляется или прекращается вовсе, растение как бы спит, дожидаясь более благоприятных условий. Поэтому причин поддерживать высокую температуру в этот период нет, потребность растений в тепле намного меньше чем в летний период.
- способные выдержать резкое изменение температур
- теплолюбивые
- любители прохладного содержания
К первой группе относят аспидистру, аукубу, кливию, монстеру, фикусы, традесканции и даже некоторые виды пальм. К любителям теплых условий зимой относятся орхидея, колеус и др. эти растения страдают от недостатка тепла и могут погибнуть, поэтому к их содержанию необходимо подходить отвественно. К третье группе относятся жасмин, цикламен, самшит и другие. Данные растения будут хорошо себя чувствовать в прохладных помещениях при средних температурах 8-12 градусов.
Обычно представители третье группы вызывают трудности, ведь в холодное время года создать прохладные условия проблематично. Да-да, ка бы смешно это не звучало, но это именно так. Люди сами по природе своей теплолюбивы, и не многие из них захотят проживать в прохладных условиях в угоду комнатных растений, да и к тому же отопление иногда жарит так что хоть окна на распашку открывай =)
Для создания прохладных условий можно ставить такие растения на подоконники, но в этом случае нужно обязательно защитить их от жара систем отопления, например отгородив защитным экраном или немного убавив обогрев
Если влияние температуры на растения и может быть разным, то резкие скачки температуры однозначно скажутся негативно. Такое часто случается, особенно зимой. Быстрые изменения температуры могут негативно сказаться на корневой системе растения, переохладить корни и листья, в результате чего растение может заболеть. Больше всего подобным перепадам подвержены растения стоящие на подоконниках, там они находятся в положении «между молотом и наковальней». С одной стороны напирает жар от батареи, а с другой холод при проветривании и замерзших стекол.
Конечно наиболее чувствительны к перепадам тропические растения, а вот кактусы стойко переносят даже сильные скачки. По природе свой кактусы находятся в условиях, когда дневная и ночная температуры могут отличаться на десятки градусов.
При проветривании комнат растения следует обязательно защищать, особенно те которые стоят на подоконнике. Для этой цели можно использовать лист картона, если защитить растения нечем — лучше убрать их подальше от окна на время проветривания.
В статье даны общие сведения, естественно, влияние температуры на растения конкретных видов могут сильно отличаться. Ознакомиться с рекомендуемыми температурами для отдельных видов растений лучше в каталоге.